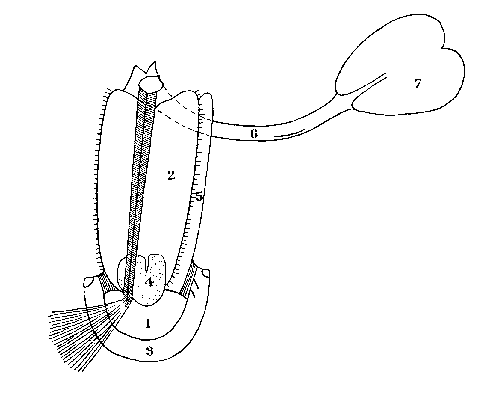
Fore-gut of the Spider Araneus diadematus.
1. Rostrum; 2. Epipharynx; 3. Labium; 4. Pharyngeal gland; 5. Hypopharynx; 6. Oesophagus;
7. Sucking stomach
General Characteristics
The
chief external feature of the Arachnida is the division of
the body into two parts, properly called the prosoma and the opisthosoma.
The former, composed of a united head and thorax, is commonly known as the
cephalothorax and the latter is also called the abdomen.
The only objection to the use of the term cephalothorax is that it
is also used for other animals in which its constitution is not the same
as in the Arachnida.
The prosoma is made up of nine segments. Some of the terga of
these prosomatic somites, and often all of them, are fused to form a
dorsal carapace, upon which the eyes, when present, are situated. Except
in the mites of the degenerate family Eriophydae, six segments of
the prosoma carry a pair of appendages. One pair only of these appendages
is pre-oral, the chelicerae. One or more of the remaining pairs
possess gnathobases, which assist in compressing and breaking up
the food particles.
In contrast to this comparative uniformity of the prosoma, there is a
much greater diversity in the appearance of the opisthosoma and of its
appendages. This part of the body is composed of twelve segments, which
are sometimes separable into a mesosoma and a metasoma,
each of six segments. It may wholly retain the segmented form or
it may be quite unsegmented, and it may have several pairs of appendages
or it may have none. In only two ways do the opisthosoma of all the
Arachnida agree - in having the genital orifice on the first somite
and in having the anus on the last somite.
The most satisfactory survey of the arachnid body may be made by taking the somites in turn.
The first is the original pre-oral somite of the ancestral annelid. In modern Arachnida, this somite is separate and distinct only in the embryo, but its existence is also demonstrated by the presence of coelomic hollows and by a nerve ganglion belonging to it. The anterior median or direct eyes are usually considered to represent its appendages.
The second somite is the one which in insects and crustaceans is called the rostral segment and whose appendages are the antennae. The absence of antennae is one of the chief negative characteristics f the Arachnida and the segment is never present in the adult. Vestiges of the appendages which belong to it have been seen during embryonic development, as by Jaworowski in the spider Trochosa.
The third somite carries the only pair of pre-oral appendages, the chelicerae. The chelicerae may be large and important weapons, as in the Solpugida, or small and relative incospicious as in the Scorpions. They are usally prehensile, but may be, as in the Spiders, unchelate.
The fourth or first post-oral
somite bears the first appendages of which the gnathobases function as
jaws. These are the pedipalpi, which may be simple tactile organs
outwardly resembling the legs, as in the Spiders, or chelate weapons of
great size, as in the Scorpions, Pedipalpi and Pseudoscorpiones.
They may also have secondary specialisations, as in the Spiders, where
they act as accessory male organs, and in the Solpugida, where they
terminate in suckers. An important fact, which does not seem to have
received much attention, is that the important and actively functional
weapon of the Arachnida is sometimes the chelicerae, sometimes the
pedipalpi, and that the more active organ increases in size at the
expense, as it were, of the other. The two pairs are never found enlarged
together.
The fifth, sixth, seventh and eight somites carry the four pair of legs in all the Arachnida. These may be all alike, as in most Scorpions and Spiders; but in the Pedipalpi, Solpugida and some of the Acari the first pair are not used for walking. They are directed forwards and carried aloft as tactile organs. In the other Arachnids the tw anterior pairs are usually directed differently from the two posterior pairs and the number of tarsal claws is not always the same. Further, the number of pair of legs which bear gnathobases is different in the different Orders.In many Opiliones all the coxa assist in masticating the food; in the Spiders this duty is performed by the maxillae or bases of the palpal coxae only, and none of the legs shares the work.
The ninth somite is of more diverse character.In the Scorpions and in the other Arachnida in which the prosoma and opisthosoma are joined across their whole breadth, the somite is not present in the adults, but its temporary existence has been observed in the embryo Scorpion. This somite persists in the Spiders and the Ricinulei, and probably also in the Pedipalpi and the Palpigradi, where it forms the waist or pedicle, uniting the two portions of the body. The somite is usually called the pregenital, since it lies next in front of the reproductive orifice. The occasional existence of both a tergite and a sternite, the lorum and plagula, above and below the pedicle, clearly indicates its nature as a seperate somite, even when much reduced in size. It forms the last somite of the prosoma when the body is divided into prosoma and opisthosoma, but is often taken as the first segment of the abdomen when the terms cephalothorax and abdomen are used.
The twelve remaining somites form the opisthosoma. They may be visible seperable into a mesosoma and a metasoma, each of six somites, as in the Scorpions, or the somites may be all alike, as in the Pseudoscorpiones and Solpugida, or they may be altogether obliterated, as in the Spiders and the Mites.
On the tenth, or first opisthosomatic, somite the reproductive organs open, and its appendages have become variously modified to form the genital operculum.
The eleventh somite is peculiar in the Scorpions. It bears the pectines, or comb-like sense-organs. In structure they resemble the gill-books of Limulus, the first pair of which belong to the same somite. In the other Orders the eleventh somite has no appendages.
Somites twelve to fifteen carry the four pairs of book-lungs of the Scorpions. The spinnerets of the Spiders belong to the fourth and fifth opisthosomatic somites, and their terminal position is due to an elongation of the third somite.
The remaining somites are seperable and recognisable in some Orders but not in all. The Scorpions retain the full number, in others the number is reduced by suppression of one or more somites.
The twenty-first and last somite carries a telson in some Orders. The Scorpions have a poison sting, the Pedipalpi a tail and the Palpigradi a jointed flagellum.
| SEGMENTATION OF THE ARACHNIDA | ||
| PROSOMA | SOMITE 1 | Pre-oral. Direct eyes |
| SOMITE 2 | (Rostral) | |
| SOMITE 3 | Chelicerae | |
| SOMITE 4 | Pedipalpi | |
| SOMITE 5 | Legs I | |
| SOMITE 6 | Legs II | |
| SOMITE 7 | Legs III | |
| SOMITE 8 | Legs IV | |
| SOMITE 9 | Chilaria, Metasoma ord Pedicle | |
| MESOMA | SOMITE 10 | Genital operculum |
| SOMITE 11 | Pectines or Book-lungs i | |
| SOMITE 12 | Book-lungs II | |
| SOMITE 13 | Spinnerets I | |
| SOMITE 14 | Spinnertes II | |
| SOMITE 15 | Book-lungs IV | |
| METASOMA | SOMITE 16-21 | No appendages |
| TELSON | Sting, tail or absent |
One of the most important
distinctions between different Orders of the Arachnida lies in their methods
of respiration, for the respiratory organs are diverse both in
nature and position.
All Arachnida are terrestrial and have book-lungs, tracheae or both.
In the Scorpions there are four
pairs of book-lungs. These take the form of small hollow sacs filled with
clusters of lamellae and opening by spiracles on the sternal plates of the
last four somites of the mesosoma.
During embryonic development the mesosoma of the Scorpion possess
vestigal limbs and it is behind the bases of four pairs of these transient
appendages that the book-lungs appear.
Similar book-lungs are found in the Pedipalpi, the Araneida and the Palpigradi.
The Palpigradi have two pairs, on the two segments of the opisthosoma following the genital segment.
The same is true of the more primitive Spiders, but in the majority the second pair have disappeared and tracheal tubes take their place, opening at a pair of spiracles on the lower side of the opisthosoma.
The five remaining Orders of the Arachnida breathe by tracheae, but these are not all found in the same situation. A summary of the facts of their distribution is best given in tabular form:
| TRACHEAL RESPIRATION IN THE ARACHNIDA | |
| 1. Acari | Eight, or fewer, spiracles in various situations - See The Acari. |
| 2. Pseudoscorpiones | Four spiracles on third and fourth opisthosomatic somites |
| 3. Opiliones | Two spiracles on second sternite |
| 4. Solpugida | Seven spiracles on second to fifth sternite |
| 5. Ricinulei | Two spiracles on prosoma above third coxae. |
Internally all the Arachnida possess a skeletal structure, known as the endosternite, in the prosoma. The existence of this is interesting, for a small endosternite of similar structure is found in Apus and in some of the decapod Crustacea, between the bases of the mandibles. In no crustacean does the endosternite attain a great size, but if the position of the Trilobites as common ancestors of the Arachnida and Crustacea be ever confirmed then the endosternite may be looked upon as persisting evidence of the relationship between these two otherwise somewhat isolated Classes.
The endosternite in Arachnida is a cartilaginous plate, slightly chitinised, lying between the nerve ganglia and the alimentary canal. To it are attached muscles from the limbs, the carapace and the pharynx. In the Scorpions, dorsal and ventral outgrowths connect it with the body wall, so that it separates the prosoma internally from the opisthosoma, and was originally described as a " diaphragm."
The internal structure of the Arachnida also includes a characteristic alimentary canal and an excretory system of coxal glands.
The mouth is a
small opening in all the terrestrial Arachnida and is seldom easy to find.
From the mouth a tubular pharynx leads to a sucking organ by the action of
which the juices of the prey are imbibed .
Fore-gut of the Spider Araneus diadematus.
1. Rostrum; 2. Epipharynx; 3. Labium; 4. Pharyngeal gland; 5. Hypopharynx;
6. Oesophagus;
7. Sucking stomach
This suctorial organ is followed
in the mesosoma by the mesenteron, but before this is reached a series of
lateral caeca diverge from the canal and run forwards into the
prosoma. These diverticula have been fully studied in spiders by
Millot , who finds that they may be divided into four types. In the first
or simple type there are but two short sacs directed forwards (see figure
below). 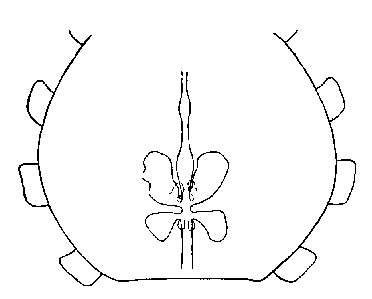
lntestinal Diverticula of Scytodes thoracica (Latreille, 1802)
Each has a more or less clearly
defined secondary portion, but the two sides are not always symmetrical.
The second or intermediate type shows three or four diverticula on each
side, by no means clearly divisible from each other and not reaching the
coxae of the legs (see figure below).
Intestinal Diverticula of Dictyna civica (Lucas,1850)
In the third or classical type,
so called because earlier workers described this type only, there are five
pairs of diverticula, one anterior and four lateral, reaching the bases of
the legs and entering the coxae for a short distance (See figure below). 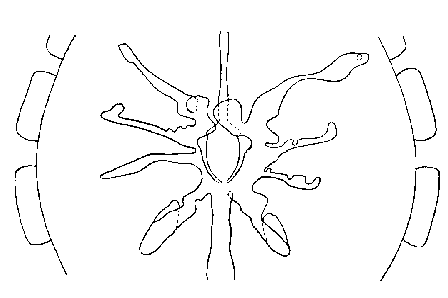
Intestinal Diverticula of Zoropsis spinimanus (Dufour,1820)
The customary statement that the
anterior diverticula meet and form a complete ring is incorrect. The
fourth or complex type differs from the third in a great development of
either the anterior or the lateral diverticula so that these divide or
branch and are so large that they quite change the appearance of the
interior of the prosoma (See figure below).
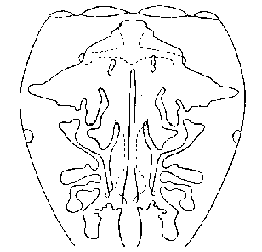
lntestinal Diverticula of Ballus chalybeius (Walckenaer, 1802)
(All above drawings after Millot)
Millot found that, although individual modifications are common, the type of arrangement is constant within the limits of a family.
Into the mesenteron open Malpighian tubes in all the Arachnida . From it there leads a complex system of branched tubules which occupy most of the opisthosoma. This great mass functions partly as a digestive gland, and partly as a reservoir. Large quantities of food-products pass into it, so that the Arachnida are well able to undergo long periods of fasting.
The coxal glands are the chief excretory organs of the Arachnida. They are nephridia, homologous with the large nephridia of segments 6 and 7 of Peripatus. In their typical form, many modifications of which are, however, found, they consist of a large excretory saccule, lined with cubical or flattened epithelium, lying outside the endosternite opposite the coxae of the first legs. This discharges its products into a convoluted tube, the labyrinth, the coils of which occupy the space from the first to the fourth coxa, or beyond. At the distal end of the labyrinth there is sometimes a swelling, regarded as a " bladder," and from this there runs forward a straight tube, the internal limb of the labyrinth, which lies inside the convolute portion and from which short exit tubes open to the exterior at small orifices behind the first and third coxae. In the Solifugae and the Palpigradi, there is an additional tube, the labyrinth sac, lined with secretory cells, between the saccule and the labyrinth. The orifice in these forms is on the pedipalpal somite. The following table shows the variations which occur among the different Orders of the Arachnida. The most variable Order is the Araneida, in which four types exist showing a progressive simplification, correlated with a corresponding increase in complexity of the silk-glands. The resemblance between the coxal glands of the Araneidae and those of the Palpigradi is interesting.
| COXAL GLANDS IN ARACHNIDA | ||||
| Saculle | Labyrinth Sac | Labyrinth | Exit | |
| SCORPIONES | In segments 5 and 6 | Absent | Many coils in segment 5 | On third legs |
| PEDIPALPI - AMBLYPIGI | In segment 3 | Absent | Very extensive coils, back to segment 6 | On first legs |
| PEDIPALPI - UROPYGI | In segments 4and 6 | Absent | Very extensive coils, back to segment 6 | On first legs |
| ARANEIDA - MYGALOMORPHAE | In segments 3 and 5 | Absent | Large, coiled, common to both saccules | On first and third legs |
| ARANEIDA - ARANEOMORPHAE | In segment 3 | Absent | Straight tube, reduced in highest to a vesicle | On first legs |
| PALPIGRADI | In segment 2 | Present extending to segment 8 | Small vesicle | On palpi |
| SOLPUGIDA | In segment 2 | Present, extending to segment 4 | Coiled, extending to segment 6 | On palpi |
NOTE.-" Segment "
in the above table corresponds to appendage i.e. the prosoma is of six
segments.
Legend: III-VI = Prosomatic
somites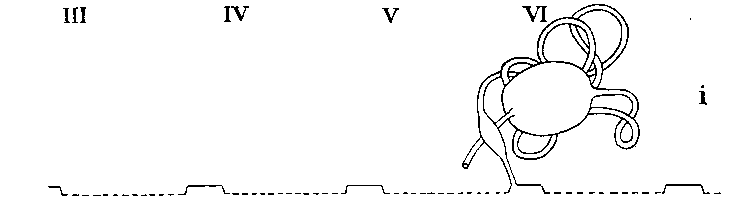
Arrangement in Scorpiones
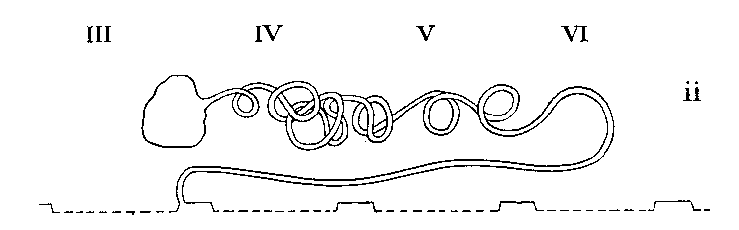
Arrangement in Tarantulini and Phrynini
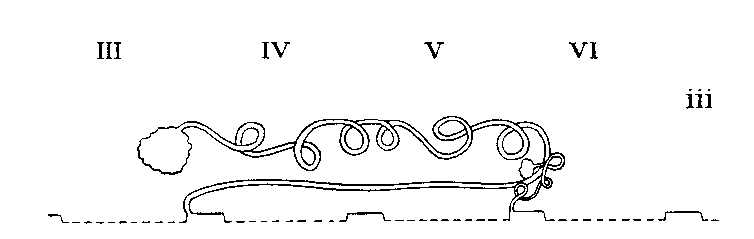
Arrangement in Charontini
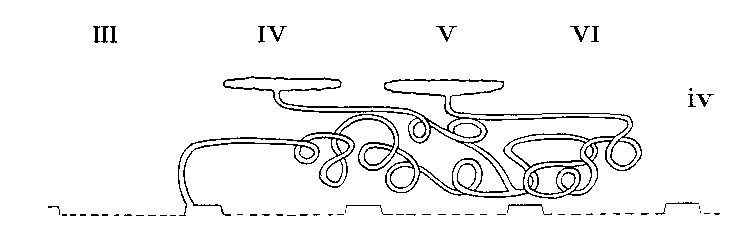
Arrangement in Thelyphonidae
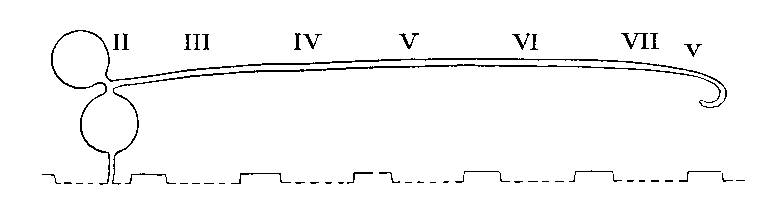
Arrangement in Palpigradi
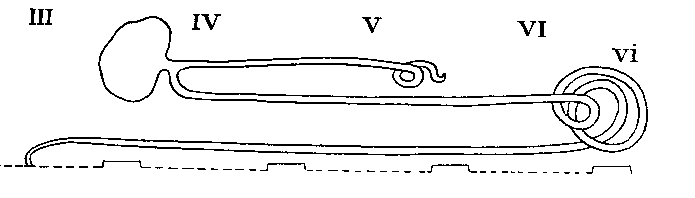
Arrangement in Solpugida
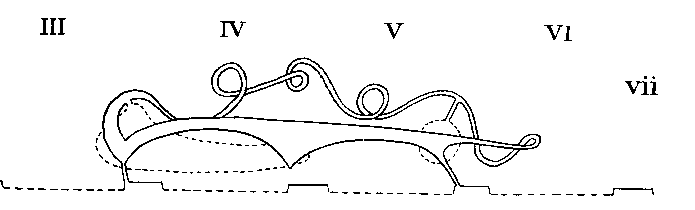
Arrangement in Mygalomorphae and Liphistiomorphae
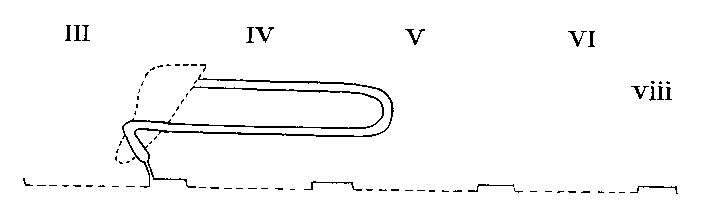
Arrangement in Dysderidae, Oonopidae, etc...
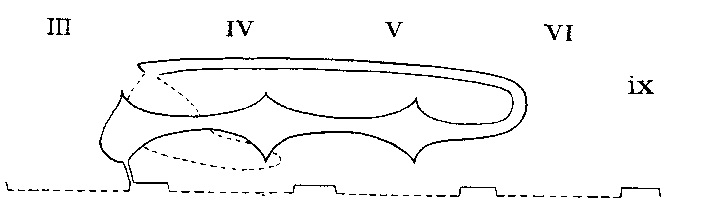
Arrangement in Lycosidae, Thomisidae, Agelenidae etc.
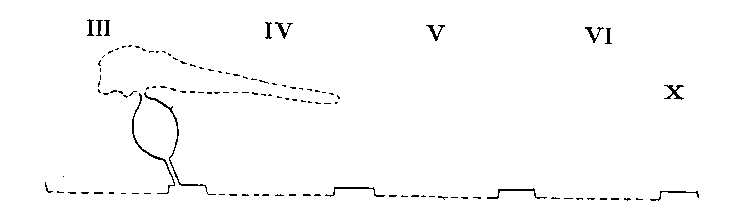
Arrangement in Argiopidae, Pholcidae, Filistatidae etc...
The difference in size between the largest and smallest of the Arachnida is not excessive. Compared with others, the Arachnida are all " small " animals, but only a few of them are excessively minute. The following table gives the lengths of a large and a small, probably the largest and smallest, species in each Order. The range would be enormously increased if extinct forms were included, for some of the Eurypterida are nearly three metres long and are the largest of all invertebrates.
| SIZE OF ARACHNIDA | ||||
| Order | Name | Size (Largest) | Name | Size (Smallest) |
| SCORPIONES | Pandinus dictator | 16cm | Microbuthus pusillus | 1,3cm |
| PEDIPALPI | Mastigoproctus giganteus | 7,5cm | Schizomus perplexus | 0,3cm |
| ARANEIDA | Theraposa blondi | 9cm | Micropholcomma bryophilum | 0,06cm |
| PALPIGRADI | Koenenia draco | 0,28cm | Koenenia grassi | 0,065cm |
| RICINULEI | Ricinoides afzelii | 1,05cm | Cryptocellus emarginatus | 0,39cm |
| PSEUDOSCORPIONES | Garypus litoralis | 0,6cm | Cheiridium museorum | 0,11cm |
| SOLPUGIDA | Galeodes caspius | 7cm | Eusimonia orthoplax | 0,8cm |
| OPILIONES | Metaphalangium propinquum | 1,2cm | Nemastoma dentigerum | 0,15cm |
| ACARI | Amblyomma clypeolatum | 3cm | Eriophyes ribis | 0,01cm |
A broad resemblance runs through the general habits of the majority of the Arachnida. They are usually animals of nocturnal activity, lying at rest during the day in their nests or holes or crouching under stones, logs or fallen leaves. The chief exceptions to this are the Scorpions, which are often active in daytime, a few Solpugida and a number of Spiders whose hunting is done by day. For most of the others sunlight inhibits movement, so that their ways are not easily determined and their habits but slightly known. In general the Arachnida are predatory. They either lie in wait for their prey and leap upon it when it approaches or they wander about and pursue it, sometimes, as in the Solpugida, at a surprising speed. There are at least two conspicuous exceptions to this. Many of the Mites are parasites and their mouth-parts are modified to form a piercing or sucking proboscis, and the Web-Spiders spin to trap and entangle insects a silk net which is almost without parallel elsewhere in the animal kingdom. One of the features of the feeding of the Arachnida is their extreme voracity. Their habits are often such that prolonged fasts are common and therefore they seem concerned to extract the last drop of moisture from each capture. The bodies of their victims are pierced or crushed by the chelicerae, mangled by the endites of the pedipalpi and legs and all liquid sucked from them by the action of the pumping pharynx within.
Among the protective adaptations
of the Arachnida colour is one of the most important. Conspicuous or
bright colours are rare, although they are found among tropical spiders
and some others that live among flowers and foliage. The majority are
soberly coloured, to achieve invisibility against a sandy or stony
background and the best of them do so with astonishing success.
Protective resemblance is also found, for there are Spiders which look
like the fruits of the plant they live on, and others that look like twigs
or thorns. Special cases of this type of protection are found, in which
the Spider makes a cocoon which resembles itself or adds a band or ribbon
of silk to its web and rests inconspicuously in the middle of this
addition. More specialised still are the zigzags of silk which tend to
distract the eye of the observer from the Spider itself.
Protection by flash-colouring also exists and is mentioned later ;
protection by mimicry is not uncommon. There are many Spiders that
very perfectly mimic ants, and others that mimic snails or beetles. One of
the most characteristic methods of protection shown by nearly all the
Arachnida is their ability to cast a limb and so to escape, purchasing
life and freedom at the price of a leg. This widespread action has been
closely studied by Dr. F. D. Wood , and she has found it necessary to
divide the circumstances in which a limb is lost into the following : (1)
Autotomy is the act of reflex self-mutilation or automatic
severance of a limb from the body. This does not exist in theArachnida,but
does exist in the Crustacea.
(2) Autospasy is the casting of a limb when pulled by some
outside agent, like the forceps of an experimenter.
(3)Autotilly is the pulling off of a limb by the animal
itself, as when an injured leg is seized in the chelicerae and thus
severed from the body.
(4) Autophagy is the act of eating a part of the body,
usually after severance from the rest.
In addition to these a fifth term, autosalizy, has been
coined by Pieron with much the same meaning as autotilly but involving,
apparently, an element of choice or decision on the part of the animal. 
The Muscles of a Spider's Leg. After F. D. Wood.
[Species Theridion tepidariorum-pro-surface of left leg.]
i. Extensor trochanteris, ii. Flexor trochanteris; iii. Flexor longus
femoris; iv. Flexor bilobatus femoris; v. Flexor bilo- batus patellae ;
vi. Flexor patellae ; vii. Protractor tibiae ; viii. Flexor tibiae : ix.
Flexor metatarsi ; x. Extensor tarsi ; xi. Flexor tarsi.
According to the view of the Arachnida this phenomenon could not be exhibited by them. It is impossible to admit that the Arachnid as it pulls on its imprisoned leg, shall come to a decision to run away on seven legs rather than perish. Dr. Wood's experiments showed conclusively that the shedding of a limb is not the result of a reflex action or of a special mechanism as had previously been believed.
It is simply the inability of
the skeletal and muscular components of the leg to resist more than a
certain force. The leg, when pulled, parts at its weakest point.
Consequently all Spiders cast their legs at the coxa-trochanter
articulation, all Opiliones at the trochanter-femur articulation and all
Pedipalpi at the patella-tibia articulation. On the other hand, in the
Scorpions and some of the Mites there is no predetermined locus of
weakness.
The musculature is evenly arranged throughout the legs and the chitin is
well provided with longitudinal fibres at all the inter-articular
membranes. These factors, combined with the lack of response on
stimulation, make autotomy and autospasy impossible and autotilly very
unlikely. Limbs which are thus sacrificed are not in general permanently
lost, but reappear. All the Arachnida undergo periodic ecdyses or
moultings of the exoskeleton, a process which is associated with
both growth and regeneration. The actual moult, which may take place in a
special rest-cocoon or in an almost unprotected situation, involves an
easy sloughing of the carapace and abdominal cuticle, a prolonged and
tedious extraction of the legs and palpi from their old cases, and a
period of rest and recovery during which the new cuticle hardens.
Bonnet has made a full study of the regeneration of lost
limbs, which reappear at ecdysis. He found that a limb is regenerated when
the interval between its autotilly and the next moult is more than
three-quarters of the time separating the two moults. If this time is
available, the length of the new limb is almost equal to the one that was
lost, and a new limb attains its full size after three moults. Hence in
order to ensure a perfect replacement, the loss must occur before the
antepenultimate moult. These generalities are as applicable to complex
organs like the pedipalpi of male spiders as to simple legs.
If a male palpus has to be regenerated in one or two moults an imperfect
and often a weird form is the result. It appears that a leg may be
regenerated as often as it is removed, and in one instance a leg was
renewed ten times. Similarly, more than one limb can be replaced at a
single moult and some of Bonnet's Spiders regenerated all eight legs
simultaneously. Another important feature in the lives of the Arachnida is
the prevalence of pre-nuptial activities when the sexes meet.
These actions, generally
described under the name of courtship, are common
throughout the animal kingdom ; they are found at least as low as the
Annelida and extend up to Man. Our present knowledge of the courtship of
Arachnida, is incomplete and is unevenly distributed among the different
Orders. The courtship of the Scorpions has been briefly described by Fabre
, who has stated that the sexes perform a kind of dance, a promenade à
deux, in which their tails are raised and entwined, until the male
grasps the female and leads her away to a sheltered spot where he digs a
burrow for her reception.
The courtship of the Pedipalpi is known only from the observations of
Fischer and of Graveley, and appears to resemble that of the Scorpions.
Fischer's description of the behaviour of Thelyphonus sepiarius
states that the female crosses her front legs which are then seized by the
chelicerae of the male. In this position the two walk slowly about, face
to face, the male now and then approaching the female more closely and
touching her with his front legs.
The same kind of behaviour is shown by the Pseudoscorpiones, a widely
quoted account of whose ways has been given by Kew , with the addition of
some degree of display by the male. The male, meeting the female face to
face, grasps her chelicerae with one or both of his own, and either
exhibits before her the peculiar ram's-horn organs he possesses, or he
shakes a palpus or his first pair of legs in a characteristic way.
The courtship of the Solpugida seems to be known only from the account
given by Heymons of Galeodes caspius. It is a much more rapid and
vigorous affair, for the male merely seizes the female with his legs and
chelicerae, grasping her body but seldom wounding her. The caresses of his
legs then so strongly affect the female that she falls into a cataleptic
or hypnotic state, in which she remains quite motionless.
There seems to be no courtship among the Opiliones and nothing is known of
the ways of the Palpigradi or of the Ricinulei.
The courtship of Spiders, on the other hand, has been very carefully
studied, first of all by Peckham in America and lately by Gerhardt in
Germany and by Bristowe and Locket in the United Kingdom. Here the manner
of courtship is varied. Male Jumping-Spiders undertake quite elaborate
dances before their females, and in these displays they are seen to
exhibit their black or coloured patches. The females carefully watch the
dance, which is ended by their joining in it. Courtship of this nature is
obviously only appreciable by Spiders with good eyesight, and the
Lycosidae or Wolf-Spiders are thus another family which behave in a
similar way. Some male Wolf-Spiders have black palpi or darkened
leg-segrnents or tufts of hair, and these decorations they wave about
before the female, raising and lowering palpi or legs as if semaphoring.
In other wandering Spiders courtship is limited to a stroking or tickling
of the female. Web-Spiders almost invariably pay court by Amending
messages along the threads of the web. The wandering male, on reaching the
outskirts of a female's web, drums on it with his palpi or plucks at it so
as to cause it to vibrate in some way which evidently does not affect the
female in the same manner as do the vibrations set up by a captured fly.
A general survey of the
courtship of the Arachnida seems to show that the result of the male's
activity is an inhibition of the female's normal food-securing reflexes.
Thus a courting male can approach a female without being slain. When near
enough he can touch her, as invariably in some way or another he does,
with the result that the necessary physical preparation for mating
commences, and in spiders with prominent epigynes actual movement of this
part has been seen. The male too, is stimulated so as to be able to exert
the considerable effort necessary for the ejaculation of spermatozoa. In
many cases this physical stimulus is so potent that the female falls into
a cataleptic state. it seems that in Spiders the details of courtship
differ for all different species, and much of the recent work has been
devoted to describing specific behaviour.
What is now needed is an intensive study of the behaviour of a single
species, so that the stimuli for each separate reflex can be perceived and
the whole process analysed into its elements as Peters has done for the
catching of web-ensnared insects.
The mating which follows courtship is differently performed in the different Orders and is often very remarkable. Only in the Scorpions and Pedipalpi is direct contact of the genital apertures found. In the Opiliones, the male organ is a very long protrusible tube, which is thrust forwards into the female oviduct as the two Harvestmen stand face to face. In the Araneida, the male organ is found in the Pedipalpi. The Spider secretes its seminal fluid either upon the ground or upon a specially spun web-the only web a mature male Spider spins-and charges its palpal organs by picking up the fluid in them. These elaborate palpi then convey the sperm to the spermathecae of the female. The position occupied by the two Spiders in the act is not always the same, but varies according to their habits and relative sizes.
In the Ricinulei, the third leg
of the male is similarly elaborated and it is assumed therefore that it
fulfils a similar function to that of the palpi of male Spiders. In the
Pseudoscorpiones and Solpugida the methods are more curious still. The
male False-Scorpion at the end of his courtship deposits a spermatophore
on the ground. This consists of an almost erect rod bearing at the top a
round globule of the vital fluid. When free from this he backs away, and
thus leads the female over it, so that the spermatophore is thrust into
her oviduct as she passes.
The male Solipugid turns the cataleptic female over and opens the genital
orifice with his chelicerae. He then deposits a mass of sperm upon the
ground, picks it up in his chelicerae and thrusts it forcibly into t he
oviduct. He then closes the aperture and presses its lips together. At
this point the catalepsy of the female vanishes and the male leaps away to
escape at his best speed. This last detail, which is shared by many other
Arachnida, shows how the inhibiting effects of the original courtship have
disappeared by the time the mating is accomplished, and the male is once
again in danger of his life from his cannibal mate.
Young Arachnida, hatch from eggs in all the Orders except the Scorpions, which are viviparous. Young Scorpions, like young Wolf-Spiders, are carried for a time on their mother's back, but maternal care is not conspicuous among the Arachnida as a whole. In some cases the young share the nest or burrow until they migrate, but direct mothering such as is found among the Theridiid Spiders, the young of which feed from their mother's mouth, is exceptional. While most Orders of the Arachnida depend for dispersal upon their own efforts, some interesting methods are adopted by others. Of these the gossamer thread of young Spiders is unquestionably the first to deserve mention. The young Spider climbs as high as possible, turns its head to the breeze and raises its abdomen. The spinnerets secrete a drop of silk which the slightest breath of air draws out into a long thread. When this streamer is pulling with sufficient buoyancy, the Spider lets go and floats away on the silk. This habit is shared by the young of several families, but only the Linyphiidae seem to be represented among the adult migrants, probably because they alone are small enough to float through the air. There is no doubt that this strange method of undirected flight, which in favourable circumstances may carry Spiders more than three hundred kilometers, is an important factor in the distribution of the Order. There are, however, many objections to the assumption that it is almost entirely responsible for the geographical distribution of Spiders as found today, not the least among which is the present scarcity of observations on the actual species known to adopt the habit.
The records of arachnologists
who have identified a dozen or a score of aeronaut species are
insufficient material - and would still be insufficient if they included
ten times as many species - for discussion of the distribution of an Order
numbering many thousands. A somewhat similar method of dispersal is found
among the very small Mites which are parasitic on fruit-trees. These
migrate in the summer, either by crawling about, or by grasping the legs
of passing Insects, or by standing erect and leaping vertically if they
feel a puff of air. The leap may then help them to be blown some distance.
The subject of aerial travel cannot be left without reference to the great
heights attained by the aeronauts. The recent work of Coad has shown that
a surprisingly numerous 4 c aerial plankton " drifts through the
upper regions of the atmosphere. His calculations point to a population,
in a column of air one square mile in area between the heights of 15 meter
and 4 kilometer, of twelve million Arthropoda in January and three times
this number in May.
Both Spiders and Mites have been found at 3 kilometer
. It is clear that the ability of wingless creatures to reach such
altitudes must greatly help their distribution. Other Arachnida achieve
dispersal by clinging to the legs of insects, like the common British Mite
Belaustium nemorum which is often to be found on the legs of
Tipulidea, and the Uropod Mites which attach themselves by a thread of
their own excrement to the bodies of beetles. The same habit is shown by
certain Pseudoscorpiones, many species of which have been found all over
the world holding to the legs of flies or of ants, or resting, sometimes
in numbers, under the elytra of beetles or the feathers of birds. This
mode of transport has been named phoresy.
The geographical
distribution of the Arachnida can be studied in detail only when
the different Orders are taken separately. When the distribution of the
Class is considered as a whole, the statement may conveniently be made
that the Arachnida are cosmopolitan, since they may be found all over the
world except in the polar regions. A rather more accurate view perceives
the Arachnida as divisible into -three groups-
(i) those which are confined to the hot tropical and very warm
sub-tropical belt;
(ii) those which spread to the limits of the temperate zones;
(iii) those which do, in fact, penetrate into the polar regions.
Scorpions, Solpugida, Ricinulei
and mygalomorph Spiders belong to the first group; Pseudoscorpiones and
Opiliones are found in the second. The third group, the polar Arachnida,
is in many ways the most interesting. There is a sharp distinction between
the Arctic and the Antarctic, due to the isolation of the Antarctic
continent and the tempestuous character of its surrounding oceans. The
first record of an Antarctic Arachnid was the discovery of the Mite, Penthaleus
belli, under moss at Cape Adare during the Southern Cross expedition
of 1899-1900. During Shackleton's Nimrod expedition (1907-9) several
species of Mites were found to be abundant in Coast Lake, near Cape Royds
on Ross Island . Skins of others were found in other lakes but none was
seen alive. A Mite, however, hatched among vegetable matter brought from
Deep Lake after the return of the expedition to England. These Ross Island
Mites were never named in the scientific reports of the expedition, but
they are of interest as being the farthest south Arachnida (77° S.).
No Spiders have been found on the continent, and the farthest south Spider
is probably either Rubrius antarticus (Karsch, 1880) from Cape
Horn or Myro frigidus Bristowe, 1931 from South Georgia (55°
S.).
Within the Arctic Circle, are
lands of comparative fertility, supporting the Esquimaux and the,
Samoyeds, with their herds, and producing flowering plants.
At least ten species of Spiders have been found in Spitzbergen, through
which the eightieth parallel of latitude passes.
The farthest north known land is the northern coast of Greenland, which
reaches latitude 83° N., only four hundred miles from the Pole.
The best work of exploration in this region was done by the Fourth Thule
Expedition led by K. Rasmussen in 1917, and their report states that "Spiders
and Earth-Mites also support life in these high latitudes". In
comparison with these records it is interesting to remember that Spiders
of the family Salticidae were found at a height of 6706 meter on Mount
Everest, in the proud position of being the highest permanent inhabitants
of the world.
After Th.Savory's Masterpiece "The Arachnids"

Deze pagina's © 1999 Gie Wyckmans & Dragon Research&Development
This pages © 1999 Gie Wyckmans & Dragon Research&Development
