Cages used in experiments on the Tropisms of Spiders
Gewoonten en Gedrag
Two alternative methods of discussing the behaviour of the Arachnida present themselves. The first is to survey the different kinds of activity - the hunting, the moulting, the spinning, the courtship, the mating and the motherhood - and to illustrate the account by descriptions of specific typical examples. is outside the scope of this book. This might be called the practical or natural history method. The alternative is the theoretical or scientific method, in which the types of action are classified and their characteristics defined. This method is probably less familiar and more valuable.
The simplest complete reaction
of the nervous system of an animal is the form of response known as a
reflex - " a neuro-muscular adjustment due to the inherited
mechanism of the nervous system."
The familiar examples of reflex action, such as the blinking of an eyelid,
are often described with an emphasis on the speed with which the whole
system carries out its function of appropriate response.
Thus there is a tendency to neglect the fact that many important reflexes
have to persist for hours at a time, an uninterrupted series of unit
actions. Important examples of this are the reflexes of posture or of
habitual attitude, the maintenance of which is their chief function. These
are called tonic reflexes. The majority of reflex actions
are quickly fatigued and quickly recover, but the tonic reflexes of
posture do not tire in this way.
On the other hand, they are the most easily dispossessed or overridden of
all, as if they paid for the continuous use of the nerve paths by an
exaggerated readiness to make way for other traffic, and in this we can
perceive their nature and function. They form a perpetual substratum of
nervous activity, maintaining the body in a state of dynamic equilibrium.
It is clearly important that this state of equilibrium shall be upset
easily, so that there shall arise that agility of response to the changing
circumstances of life which produces efficiency and successful activity in
the living animal. A very large proportion of the life of an Arachnid is
spent in waiting, and during these long periods of inactivity the tonic
reflexes are in sole possession of the nervous system, supporting the body
on its eight legs against the unceasing gravitational force.
For animals like the Harvestman Liobunum rotundum, with its tiny body and eight long symmetrically arranged legs, the tensions in the muscles of all legs are probably almost equal, and for Spiders like the Linyphiidae which hang inverted below their hammock-like webs, the tensions are probably not very unequal.
It can scarcely be a coincidence that these types are particularly sensitive to surrounding disturbances. In the common Garden-Spider, which hangs head-downwards in its lovely orb-web, the weight must clearly be disproportionately supported by the two hind pairs of legs and the same is true of almost any Arachnid that stands on the ground. The heavy opisthosomatic region has no legs of its own and the greater part of its support must come from the posterior pair of thoracic legs. It should not be surprising, therefore, that when the equilibrium of the tonic reflexes is upset, the readiest response is a flexion of the femora, so that the patellas tend to meet over the middle of the animal, and especially a flexion of the forelegs. This simple automatic response, which may be called the flexor reflex, is easily witnessed whenever a Spider is, as we should say, frightened, and several valuable consequences follow from its operation.
One of these is the flash-colouring
method of protection, which most naturalists would associate with
the Tree-Frogs of tropical forests, but which is also well shown by
several native Spiders.
A good example is Segestria senoculata, a common species with
diamond-shaped marks on its femora and bright tawny femora. As it runs,
the moving femora are conspicuous, but when it suddenly stops and draws in
its legs this brightness vanishes and the observer finds the Spider
invisible. Without any help from flash colours, the flexor reflex assists
in producing invisibility in other Spiders.
If on the beach in certain parts of the coast the heaps of dried seaweed
are turned over, there runs out a sombrely-coloured active Spider, Philodromus
fallax.
As it runs it may easily be mistaken for a grain of sand, rather bigger
than the average, rolling down a slope, and, in fact, such grains of sand
are often mistaken for Spiders. But there is this difference. When the
pellet of sand comes to rest it can be seen ; when the Spider stops moving
it is invisible. It does not bury itself nor leap suddenly aside; it
simply stops and flexes its legs. And it is gone. The writer is not
short-sighted, but on the Norfolk coast he has stared straight at these
Spiders and been quite unable to see them until they have moved.
This leads naturally to the
obvious and probably the most important of all the consequences of the
reflex, the action metaphorically described as " feigning death and
less inaccurately as the cataleptic reflex. In many
Arachnida a sudden disturbance causes them to draw in their legs and fall
motionless, and in some the caresses of the opposite sex produce the same
condition. Various considerations make it clear that the animal is not in
a true state of catalepsis.
Often after first falling motionless it will re-arrange its legs slightly,
and in some species a periodical tremor runs through the limbs. In
addition to these movements, which would not be seen in true catalepsy,
there is not the complete insensibility which would be expected. If the
motionless Spider be gently touched it will often get up and run away. If
the Spider is lying low at the end of a thread attached to its web and the
web be touched with a vibrating tuning-fork, the Spider at once awakes and
returns to its web. The whole action is really nothing but a sustaining of
the flexor reflex and the more closely it is studied, the more clearly it
shows the characteristics of reflexes in general.
Many reflexes continue to discharge after the stimulus ceases, and in the "
cataleptic " Arachnid we are witnessing the retention of the position
produced by the flexion of the femoral muscles. All reflexes, however, are
subject to fatigue which causes them to give up possession of the
nerve-paths in favour of some other impulse, previously inhibited. Thus
the quiescent Spider awakes and a response to a probably new situation
occurs. In several families of Spiders the flexor reflex operates with a
different result.
The common House-Spiders,
Tegenaria, are familiar examples of Spiders which rest in the corner of a
sheet web. Their forelegs are outstretched and their claws grasp the silk
so that often the sheet can be seen to be drawn up into small cones. If an
insect brush against the web, the legs are jerked inwards, plucking
sharply at the sheet ; and the result is not invisibility for the Spider
but a further entanglement of the insect.
Orb-web Spiders perform exactly the same action, the web shakes and the
Spider plucks at it. Very often the Spider then turns about and repeats
the jerking in another direction, an action which, despite its intelligent
appearance is only a consequence of unequal tensions in the threads of the
web, The inequality may be due to the recently added weight of the captive
and it is this which turns the Spider round, and not an attempt to
discover whereabouts in the web the arrival has landed.
Like beauty, which lies in the eye of the beholder, purposiveness lies in
his interpretation of what he beholds.
A consideration of a second reflex action furnishes support for this
view. If a reflex is found to have no manifest result or no meaning in an
anthropomorphic sense, the observer refrains from imputing purpose to the
animal and is content to describe the action itself as a mystery.
Spiders often exhibit one such action. If the flexor muscles raise the
femora while the extensor muscles straighten the patella, the result will
be that the Spider lifts its leg into the air, stretched up at right
angles to its body. This is most often seen in orb-web Spiders, for which
it constitutes the normal reaction to a sudden noise. A whistle, a cough
or the bark of a dog near a full-grown Araneus usually makes it shoot out
its forelegs, as if reaching towards the origin of the sound. The
anthropomorphic description of this action would be that the spider is
listening, but whatever it is that the Spider hears it does not proceed to
any further action, and the idea that it can hear better with its toes in
the air is too foolish to be suggested.
Moreover, other Spiders perform the same action in quite different
circumstances. Some female Spiders in charge of their egg-cocoons assume
this position when driving off intruders, others similarly repel the
advances of an unwelcome male. Many Spiders raise their forelegs in
courtship, others do so when teased or threatened. It seems reasonable to
interpret the otherwise meaningless act by suggesting that it is elicited
by a different kind of disturbance from that which produces complete
flexion. The femora are not drawn back so far, the tibiae are not folded
against them, the posterior pairs of legs are not moved. Still another
consequence of the flexor reflex remains to be considered.
In the performance of many
mammalian reflexes there is a characteristic known as the
refractory phase. This is the periodic recurrence of a condition
of inexcitability of the flexor muscles, so that the limb is momentarily
straightened by the extensors, and the result is a rhythmically repeated
motion as in scratching.
Arachnida do not scratch themselves, but some of them in certain
circumstances will show a rhythmic repetition of the flexor reflex, which
again has valuable results. Two very common Spiders, Araneus
diadematus and Pholcus phalangioides possess the habit of
rapidly shaking their webs so that they themselves, usually conspicuous in
the centre, become blurred and indistinct.
This shaking is achieved by vigorous contractions of the femoral muscles,
suitably timed, but even if the stimulus continue the reflex will cease.
Reflexes can be stopped either by fatigue or by inhibition from another
reflex. Under inhibition the reflex fades out with no change in the
frequency or extent of the beats, and this is seen if one touch the
oscillating Spider, causing it to drop at once. Under fatigue the actions
continue with a slower rhythm and this is seen if the Spider can be
forced, as by gently blowing on it, to continue its vibratory movements.
In this account of reflex actions it is seen that the well-known reflexes of Mammals are of the same nature as the reflexes of Arachnida, and that these reflexes are responsible for many of the essential actions of an Arachnid's life. Even the familiar conditioned reflexes can be paralleled and the work of Dr. Pierre Bonnet of Toulouse provides an instance. During his study of the Spider Dolomedes, it was his custom to keep many specimens, each in a covered glass jar. These captives were fed by presenting to them a fly held by a wing in a pair of forceps, upon which the Spiders unhesitatingly leapt. After a while, the act of raising the cover-glass caused the Spiders to turn and leap forward, although the fly had not entered the jar. The Spider's pounce, originally a response to the buzzing of the fly, became conditioned to the opening of the jar. Once again it is true that the behaviour which wears so striking an appearance of consciousness and purposive striving is but a balance of physiological processes, stimulation and inhibition, which take place in a body or biochemical system of alternating equilibrium-states.
A very extensive group of
actions which cover a large proportion of an Arachnid's activities are
known as tropisms. A tropism is a directed reflex,
concerned with bodily orientation in response to a particular type of
stimulus, so that symmetrically placed sense-organs receive equally
intense stimulation.
Light, water and gravity, responsible for phototropism or heliotropism,
for hydrotropism and for geotropism, are the most familiar external causes
of tropistic behaviour and Arachnida provide good examples of them all.
The effect of light is often very marked, and ordinarily its most obvious influence is the way in which it deadens the irritability of organisms, many of which are quiet all day and are then seized with a strange restlessness and a readiness of response during the night. The most familiar Spiders spin their webs in the evening, and Harvestmen provide an even better example. When living in captivity they are so still during the day that to a casual observer they seem to be dead, while at night they are active and eat the food which has been supplied to them and which they have neglected while the sun shone. On the other hand Wolf-Spiders and Jumping-Spiders are generally busy in the sunshine, but disappear as if by magic in response to a passing cloud.
Reactions to gravity are also common among Arachnida, and many Spiders when kept in captivity invariably climb to the highest part of the cage. Several Hunting-Spiders become geonegative and climb into the undergrowth when they are mature, while Harvestmen hunt on the ground at night but generally climb up trees or houses for rest during the day. The most pronounced example of negative geotropism is shown by the young Spiders which are dispersed by an aerial flight on a gossamer thread, for the little Spider always climbs to some height before it sets sail.
But many other kinds of tropism besides these three can be detected in the study of the habits of small animals. Stereotropism is a reaction to bodily contact, hygrotropism to water vapour, chemotropism to different chemical compounds, thermotropism to heat, vibrotropism to vibrations, galvanotropism to electric potential, rheotropism to visual images and anemotropism to wind pressure.
The Arachnida can provide good examples of all these types of tropistic behaviour. Anemotropism is a term used by Wheeler to describe the orientation of insects to the direction of the wind, so that they fly with equal pressure on their wing surfaces. As far as the Arachnida are concerned, an especially good example of this not very common tropism is provided by young Spiders, during their migration on threads of gossamer, mentioned above. When they have climbed to the summit of their ascent, they turn head to wind before raising the abdomen and secreting the silk thread and this turning is directed solely by the wind.
Rheotropism is
in part an allied phenomenon in which the pressure is supplied by water
and thus is common in many animals which live in rivers and running
streams. It prevents their being washed out to sea. Just as geotropism may
be effective through the semi-circular canals, the eyes or the pressure
upon the feet, or normally through all these, so rheotropism has
apparently a dual mechanism, effective through lateral sense-organs
sensitive to touch and also through retinal images. For the latter there
is an apparent tendency to keep visual images at a fixed spot on the
retina.
Fish show this well and swim upwards if an object beside their aquarium
move downwards. Spiders show the same tropism. If the hand is moved over a
running Spider from behind, the Spider can be made to turn and run in the
opposite direction.
Opiliones, Harvestmen, do not react to a hand above them since their eyes
look outwards, not upwards. But if the hands are placed one on each side
of the animal and are moved up and down, the Harvestman's body rises and
falls on its long legs in unison, with the most ludicrous appearance
possible.
Stereotropism,
a tendency to force the body into corners and crevices, is well developed
among most of the Arachnida, which are impelled by its insistence, aided
by negative phototropism, to hide themselves in safety during the day.
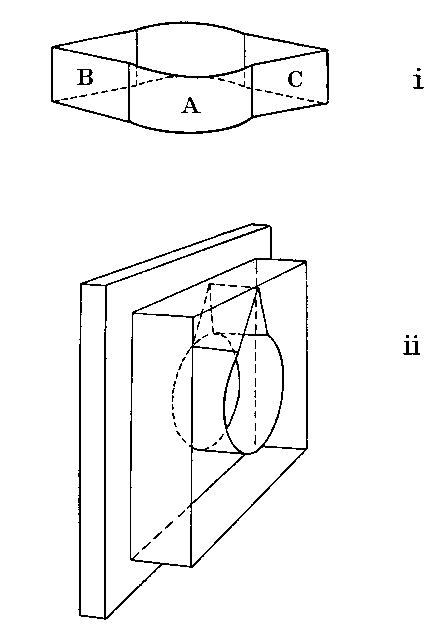
Spiders may be used to give most interesting results in experiments on
this tropism by putting them into a cage (see figure below) of cylindrical
form with two angles, one made of glass (B) and the other of cardboard
(C). When a Spider is put into this cage it has the chance of remaining in
the circle (A) or going into either the dark or the light crack, and the
cage therefore not only detects stereotropic tendencies but compares
stereotropism with either positive or negative phototropism.
Cages used in experiments on the Tropisms of Spiders
It is not to be expected that many Spiders will be stereo-neutral, but one at least, Pisaura mirabilis, was found never to attempt to force itself into either crevice. When a Spider, such as Amaurobius ferox, whose normal habit is to live in a dark corner is put into the cage, there is no delay in its entering the crevice, but it will not enter the glass angle if the dark one is available. In a cage with a glass angle only it will enter the angle instead of remaining in the circle, thus showing that its stereotropism is stronger than its negative phototropism. In the form of the cage shown in the figure, ii, , there is one crevice only and the floor is vertical. It is pivoted to a board by a pin through the centre of the circle, about which it can be revolved, so that it can be used to compare the strengths of stereotropism and geotropism. The Spider Steatoda bipunctata is stereopositive and very markedly geonegative. It invariably climbs into the crevice when this is pointing upwards, but it simply will not go down into it. Thus its negative geotropism is stronger than its stereotropism. Its phototropism is feeble, and the substitution of a glass crevice for a dark one does not change the result.
Hydrotropism, or turning towards water, is well shown by the Arachnida, many of which are habitually thirsty creatures. If a drop of water be cautiously lowered with a pipette into a cage containing a thirsty Spider, there is at first no response. The drop begins to evaporate and the vapour diffuses across the cage and so reaches the Spider. Then, after a time, there is a response ; the Spider moves its palpi, as if testing the air. Finally, as the concentration of water vapour in its neighbourhood rises, it moves towards and drinks the drop of water. This experiment shows that the thirsty Spider does not move about seeking water, as a man would but that, if its internal organs are sufficiently desiccated, its forced response is such that it is led towards the liquid it requires.
Hygrotropism, a term apparently first introduced by Th. Savory, differs from hydrotropism since it is solely a reaction to atmospheric water vapour. Water vapour is an extremely active component of an invertebrate's environment, and many creatures, and especially the Arachnida, are sensitive to hygroscopic change. This tropism is responsible for many of the peculiarities in the localisation of Spiders to small areas within their total range.
Vibrotropism is
the response of an animal to vibratory disturbance of its surroundings and
among several examples Web-Spiders are undoubtedly the animals in whose
lives it plays the greatest part. By it they are guided directly to the
source from which the vibrations of their webs originate. The Spiders that
run on the surface of water respond in the same way to the ripples
produced either by struggling insects or by the prong of a vibrating
tuning-fork. It can readily be seen that this tropism is a direct
consequence of the flexor reflex. Let it be supposed that the Spider is
standing with all eight legs on the surface, as shown in the following
figure:
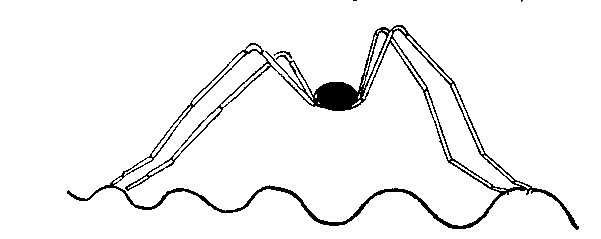
Diagram to illustrate the Mechanism of Vibrotropism.
The ripples passing under the
right legs will have a smaller amplitude than those passing under the left
legs, and thus there is a more intense stimulation of the right legs,
followed by a greater degree of flexion of the femoral muscles. The
consequence of this forced movement is that the extended left legs exert a
turning effect relatively to the less extended right legs so that as the
spider runs it is automatically directed towards the origin of the
disturbance.
The number of recognised tropisms indicates an important aspect of animal
behaviour, for if an organism be subject to ten or a dozen modes of
tropistic response to external conditions,'it is seldom that in natural
circumstances a single tropism can act alone, holding the animal as it
were at its sole mercy. Thus we are led at once to consider an interplay
of tropisms, and to regard the organism as a particle subjected to a
number of possibly contending forces and obedient to their algebraic sum.
In this very fact there is always the risk of a facile statement that some
tropism is involved without any proof of the truly tropistic nature of the
actions.
By constant usage one may commit the peculiar type of fallacy known as a
reification of words, and come to regard a tropism as if it were a thing
which, like a gland, may or may not be found in an animal.
But a tropism is not a thing, it is a way in which organisms behave. The
body reacts to changes in its environment by secretion or by movement.
When movement has occurred its direction is governed by tropistic
principles. An examination of the distribution of the Arachnida shows that
they are found in situations where the conditions are most nearly constant
; they are to be found in ditches, under stones, leaves and logs of wood,
and in caves. This apparently ascribes the selecting agency to the
activity of the animal. But the boot is on the other leg : the real
selector is the environment.
It is not drought or flood, cold or warmth that disturbs the
invertebrate, but the change from one to the other. Should the conditions
alter so as to overstep the various thresholds, the animal is affected and
must respond. Thus it is inexorably moved on, as if by a merciless
policeman, until it reaches a station in which it can remain. Here it is a
prisoner, until further change releases it. This conception of an animal,
devoid of behaviour in a constant environment, is, unfortunately, contrary
to ordinary ideas and ordinary experience. But it is a fact that ordinary
experience is seldom derived from the study of animals in their normal
surroundings.
A naturalist usually meets small animals just after he has disturbed them.
He belabours the hedges and ploughs up the leaves or at least goes
crashing through their neighbourhood, his titanic feet shaking their
world, his shadow obscuring the sun, his scent lingering behind him. Only
a few observers realise how large a proportion of an animal's time is
spent doing nothing.
Here is the true place of the study of tropisms in animal behaviour. Any
particular tropism can, of course, be understood only after experimental
study under laboratory conditions, yet in nature animals exhibit tropistic
behaviour although no mechanistic biologist stands by to measure their
angles of deviation. The customary description of animal behaviour as "
instinctive " demands further consideration.
Instinctive acts,
like reflexes, do not have to be learnt and they are adaptive in the sense
that they tend to the preservation of the animal and of the race when they
are normally performed in normal circumstances, but when circumstances
change it is often found that the animal neither modifies nor reverses its
instinctive procedure, continuing its course unchecked with useless or
even fatal results. But, unlike reflexes, instinctive acts demand an
intact nervous system, and they are often directed towards a relatively
distant goal. Many of the deeds that animals perform on behalf of their
young will bear fruit only in the future, while reflexes are always
concerned with affairs of the moment. Further, instinctive actions are
sometimes capable of modification. All these characteristics of
instinctive actions are well illustrated by the Arachnida.
The spinning of the Spider's web, the building of the False-Scorpion's
nest, the making of an egg-cocoon, the courtship of Scorpions, Solipugids,
Spiders and False-Scorpions and the elaborate behaviour involved in the
life-histories of Mites and Ticks are all instances of typically
instinctive actions.
As long as we are studying the
Arachnida we are dealing with instinctive actions in which their castiron
routine is well displayed. The well-known experiments of Fabre and the
even more critical studies of Hingston have uniformly emphasised this
fact, that instinctive behaviour may develop in an irreversible sequence
and is usually quite unable to deal with any situation outside the
ordinary. This is a description of the nature of instinct and not a
criticism of the animal's ability, for the instinctive actions normally
suffice for the animal's survival.
The young Spider spins its first web quite perfectly - and a year or so
later it may spin its last. The last will be no better and no worse than
the first, it will not be spun more quickly or in a better place. The
Spider has not profited from past experiences, and so has robbed us of all
the evidence we might have had that the experiences were conscious ones.
All its life it has just spun - instinctively, irresistibly, irrationally
- for it has no consciousness, it has no mind. Such instinctive
behaviour is, in general, largely governed by internal physiological
conditions, just as tropistic behaviour is governed by external physical
conditions.
The habits of the Water-Wolf
Spider, Pirata piraticus, illustrate this particularly well. These
Spiders spin a tube-like retreat leading to the water's edge and, as
mentioned above, react to'water ripples as Web-Spiders react to the
vibrations of their snares. When the female has laid her first cocoon she
remains in the tube but neglects to repair it. It is not that she never
spins, for at regular intervals she detaches the cocoon from her
spinnerets, holds it in her third pair of legs and turns it about while
adding more silk to the outside. She will then lay it aside for a short
while and add a little silk to her home, but her instincts are those of a
mother, not of a builder, and the tube soon becomes a wreck. By the time
that the first cocoon has hatched and the young have dispersed, the
ovaries will be maturing for a second family, and shortly before this
cocoon is laid, the Spider reverts to her maiden habits and spins a new
and perfect home. This process of alternating neglect and replacement is
repeated between the second and third cocoons, after which the summer is
almost over and the winter torpor is approaching.
When a number of these Spiders are being kept at the same time, the
influence of.approaching and receding maternity upon their normal
instincts is particularly striking.
Thus one important aspect of the nature of instinct begins to appear.
To the older naturalists this presented such great difficulties that they
were usually forced to be content with description and metaphor. Thus they
spoke of instinct as " racial habit " or as "inherited
memory" or as " lapsed intelligence", but these
were mere verbalisms and no progress results from sheltering behind words.
Some advance has now been made from that position, so that we now look upon instinctive actions as being reflex acts, following one another in predetermined succession, coming often under the influence of external circumstances, so that a symmetry of activity is produced, and often also under the influence of internal conditions, so that new types of activity constantly appear. This union of external and internal factors and the realisation of the nature of the latter is important.
Instincts are something more
than reflexes, but this something " is of a material nature, secreted
by glands it acts as a chemical compound distributed by the blood. Thus it
becomes at once subject to direct study, and mere difficulties of
technique are the obstacles to a full knowledge of the nature of every
hormone, of the way in which it is produced and the physico-chemical
nature of its mode of action.
An exceptionally fine analysis of an instinctive action in this way has
been made by Peters, in his study of the capture of insects by the common
Spider Araneus diadematus. He concludes that a series of stimuli,
each followed by a characteristic reaction, produces the habit and that a
psychical factor nowhere encroaches in the chain. The capture of a fly is
thus analysed by Peters into five separate elements, as follows :
i. Movement towards Prey. The Spider is guided by the
vibrations of the web, or if the prey does not move, by the change in
tension of the threads. The stimulus for this forward movement is the
sudden movement of the web.
ii. The Reflex of the Long Bite. The stimulus for this is
the vibratory movements of a struggling fly : stationary objects are not
bitten in this way.
iii. The Wrapping or Enshrouding Reflex. This familiar
act is a compound reflex legs and spinnerets being involved, the stimuli
for which are both chemical and tactile. These stimuli are received when
the prey is touched with the forelegs and palpi, and when it is bitten.
iv. The Reflex of the Short Bite. This follows the
wrapping of the prey and the stimulus is a tactile one received from the
silk threads with which it is covered.
v. The Carrying Reflex. The wrapped prey is carried to the
Spider's waitingplace ; the stimulus is the chemical one received from the
short bite.
Peters succeeded in calling out some of these reactions isolated from the process of capture. " If," he writes, " one holds to the Spider in forceps a struggling fly, the insect is bitten (long bite). If one cautiously touch the palpi with a freshly killed fly, it is immediately wrapped up. If one use an already wrapped fly, it is bitten (short bite)." In later work he has further investigated the fifth element, the " carrying reflex." It is well known that this spider sometimes carries away its booty in its chelicerae, but sometimes it attaches the wrapped packet to its spinnerets. Peters has shown that the weight of the prey determines the method of transport, relatively heavy objects of about 80 mg. being carried on a thread with the help of the hind legs, while lighter ones of about 10 mg. and the same dimensions are carried in the chelicerae. His papers contain many other valuable observations, of great interest to those who believe in his method of studying and analysing the behaviour-patterns of Spiders.
After Th.Savory's Masterpiece "The Arachnids"

Deze pagina's © 1999 Gie Wyckmans & Dragon Research&Development
This pages © 1999 Gie Wyckmans & Dragon Research&Development
